



بخشی از مقاله
مقدمه
در راسته آلالهسانان (Ranunculales) تیرة آلالهایان موقعیت فیلوژنتیک برجستهاي داشته و به نظر میرسد پایه و اساس یودایکتها باشد. این تیره یک تیرة بزرگ با پراکنش جهانی
است (Soltis et al., 2005; Simpson, 2006; Heywood
.et al., 2007) این تیره تنوع وسیعی در ویژگیهاي ریختی به-ویژه در نوع میوه و سازمانیابی گل نشان میدهد و به همین دلیل طبقهبنديهاي متعددي براساس ویژگیهاي ریختشناختی
(Hutchinson, 1923; Janchen, 1949; Tamura, 1995)، ویژگیهاي ملکولی (Jensen et al., 1995; Ro et
al., 1997) و ترکیبی از ویژگیهاي ریختشناختی و ملکولی انجام شده است .(Wang et al., 2005) این تیره بهکوشش (1995) Tamura به سه زیر تیره و یازده طایفه تقسیم شده است. این تقسیمبندي براساس عدد کروموزومی، برچه و نوع میوه انجام شده است اما در این مورد هنوز اتفاق نظر وجود ندارد.
گیاهان تیرة آلالهایان شامل ویژگیهاي متعدد ابتدایی و پیشرفته ریختشناختی و رویانشناختی است: گلهاي منظم یا نامنظم دو یا تکجنسی، برچههاي آزاد یا پیوسته، فولیکولهاي چندبرچه-اي یا فندقههاي تکبرچهاي، تخمکهاي کم یا پرخورش تک یا دوپوسته اي، تکوین کیسه رویانی از نوع Polygonum یا Allium و آنتیپدهاي پایدار یا ناپایدار ازآنجمله است، اما از یک گونه به گونه دیگر تنوع قابل توجهی در ویژگیهاي فوق
وجود دارد (Vijayaraghavan & Bhandari, 1970;
.Johri et al., 1992) وجود پلیپلوئیدي مکرراً در انواع سلولهاي کیسه رویانی نظیر آنتیپودها، سینرژیدها، آندوسپرم و
سوسپانسور گزارش شده است (Kozierradzka-Kiszkurno
& Bohdanwicz, 2003; Greilhuber et al., 2005, . Ghahremaninejad et al., 2013)
وجود پلیپلوئیدي مکرراً در انواع سلولهاي کیسه رویانی نظیر آنتیپودها، سینرژیدها، آندوسپرم و سوسپانسور گزارش شده
است (Kozierradzka- Kiszkurno & Bohdanwicz,
2003; Greilhuber et al., 2005; Ghahremaninejad et al., 2013) و ارتباط آشکاري بین درجه پلیپلوئیدي این
سلولها و طول مدت زمان زندگی آنها وجود دارد که نقش
68/68 Nova Biologica Reperta 1 (2): 67-76 (2015)
تغذیهاي آنها را پیشنهاد میکند .(Raghavan, 1997) وقوع آندوپلوئیدي در اعضاي این خانواده بسیار شاخص است
(Turala-Szybowska et al., 1981; Turala-.Szybowska & Wolanska, 1989)
هدف این مقاله مطالعه مراحل تکوین تخمک و کیسه رویانی در گونه C. orientalis بهمنظور توسعه دانش رویانشناسی این تیره است. براساس مطالعات مرجعشناختی ما این جنس و گونه ازنظر تکوینی تحت مطالعه قرار نگرفته است. بهعلاوه، آشکار-ساختن خصوصیات تکوینی تخمک وگامتوفیت ماده سبب فهم بهتري از مسائل آرایهشناختی تیرة آلاله میشود.
مواد و روشها
گلها و جوانههاي گونه تحت مطالعه در اوایل تابستان از زیستگاههاي طبیعی دراطراف شهر همدان جمعآوري شد. گلها و غنچهها در مراحل مختلف تکوین جهت تثبیت در
(Formalin, Acetic acid, Ethanol) FAA70 قرار داده شد، سپس ذخیرهسازي در الکل 70 درصد صورت گرفت. پساز قالبگیري، برشگیري نمونهها در قالبهاي پارافین با میکروتوم (Microm DC, Dideh Sabz, Iran) با ضخامت 7-10μm صورت گرفت .(Xue & Li, 2005) رنگآمیزي برشهاي تهیهشده پساز پارافینزدایی و آبدهی با روش رنگ-آمیزي مضاعف هسته و سیتوپلاسم توسط هماتوکسیلین و ائوزین صورت گرفت .(Yeung, 1984) نمونهها با میکروسکوپ نوري (Germany) Zeiss Axiostar Plus
بررسی شد و عکسبرداري به کمک دوربین دیجیتال مدل
(Japan) Canon G11 انجام گرفت.
نتایج
تشکیل مادگی و تخمک
تخمکها به صورت برجستگیهاي کوچک کموبیش کروي تا بیضیشکل در جدار تخمدان پدیدار میشود (شکل .(1A تخمدان از نوع فوقانی و دوخانهاي است و هر حفره دو ردیف تخمکی دارد (شکل .(1B برچه شامل تعداد زیادي تخمک است. تخمکها در تخمدان به طور واژگون و آویخته در دو
یافتههاي نوین در علوم زیستی جلد 1، شمارة 67-76 :2
ستون قرار دارند. جایگزینی تخمکها در تخمدان از نوع جداري است. پریموردیوم تخمک داراي 4- 7 ردیف سلولی است (شکل .(1A در C.orientalis چندین سلول آرکئوسپوري وجود دارد (شکل (1C و تنها یکی از این سلولها به طور مستقیم به مگاسپوروسیت تمایز مییابد. سلول مادر مگاسپور در نمونه تحت مطالعه، در لایه چهارم است (شکل .(1D پریموردیوم تخمک خمیدگی را از همان مراحل اولیه آغاز میکند. هنگامی که پریموردیوم تخمکی کشیده میشود، در بخش میانی آن ابتدا پوسته خارجی بنیانگذاري میشود (شکل .(1F بعد از مدتی، با توسعه پوسته خارجی، پوسته درونی در بخش درونی پوسته خارجی تشکیل میشود (شکل .(1 E در نمونه مورد مطالعه، تخمک از نوع دوپوششی، پرخورش و واژگون است (شکل .(3B میکروپیل بوسیله پوسته داخلی که متشکل از 2 لایه است شکل میگیرد و از نوع خطی صاف است (شکلهاي 4B و .(4A پوسته خارجی ضخیم و متشکل از 4 تا 5 لایه سلولی است. از درونیترین لایه پوسته درونی، لایه اندوتلیوم تشکیل میشود. سلولهاي این لایه به صورت شعاعی امتدادیافته و داراي هسته کاملاً واضح و سیتوپلاسمی متراکم هستند (شکلهاي 4B و .(4A در C. orientalis تشکیل مادگی پساز پرچمها صورت میگیرد. مادگی تکبرچه و داراي پایک و کرك است (شکلهاي 4F و .(4G
مگاسپورزایی و نمو گامتوفیت ماده
سلول مادر مگاسپور را چندین لایه از سلولهاي خورش احاطه میکنند، بنابراین تخمک از نوع پرخورش است. سلول مادر مگاسپور ازنظر اندازه رشد میکند و با حجم زیاد، هسته درشت، سیتوپلاسم متراکم و شکل چندوجهی خود از سایر سلولهاي خورش قابل تشخیص است (شکل .(1D حضور کلاهک خورشی از این مرحله دیده میشود که بین دو تا چهار لایه است. مگاسپوروسیت با انجام میوز مگاسپورها را تشکیل می-دهد که ضمن رشد گامتوفیت ماده یا کیسه رویانی را تولید میکنند. اولین تقسیم میوزي مگاسپوروسیت، موجب تشکیل
/69 Nova Biologica Reperta 1 (2): 67-76 (2015)
69
دو هسته هاپلوئید (شکل(1E و پساز سیتوکینز دو سلول دیاد هاپلوئید میشود (شکل.(1F در نمونه لامهاي تهیهشده از مراحل تکوین تخمک در این گیاه هم دیاد طولی (شکل (1F و هم دیاد عرضی مشاهده شد (شکل .(1G دومین تقسیم میوزي موجب تشکیل چهار سلول تتراد میشود. انواع تترادها را در این نمونه میتوان مشاهده کرد. ازجمله نوع خطی طولی، خطی عرضی 4) سلول کنار هم)، T شکل و چهار وجهی (شکلهاي .(2A-2D از چهار سلول حاصل از میوز، سه سلول تحلیل می-روند و فقط یک سلول (مگاسپور عملکردي) باقی میماند. در این نمونه، موقعیت سلول باقیمانده یا مگاسپور عملکردي سفتی (میکروپیلی) است.
در C.orientalis وقتی میوز با تشکیل تتراد به پایان میرسد، پوسته تخمکحدوداً 3/4 جسم تخمک را میپوشاند (شکل .(3A مگاسپور عملکردي سه تقسیم میتوزي متوالی انجام می-دهد و درنهایت کیسه رویانی هشت هستهاي و هفتسلولی را به وجود میآورد. طی تشکیل کیسه رویانی ساختارهاي هیپوستاز و پوستامنت شکل میگیرند (شکل .(4A سه سلول در قطب مجاور سفت استقرار مییابند و دستگاه تخمزا را به وجود میآورند. دستگاه تخمزا شامل یک سلول تخمزاي بزرگ و گلابی شکل است و به واسطه داشتن یک هسته درشت و یک واکوئل بزرگ قابل تشخیص است، و دو سلول کناري آن "سینرژیدها" است. در قطب بنی، سلولهاي آنتیپود به وجود میآید که آنها نیز آرایشی مثلثیشکل دارند. در نمونه مورد-بررسی این سلولها غولپیکر بوده، از قدرت رنگپذیري بالایی برخوردارند و به سمت مرکز کیسه رویانی مهاجرت میکنند. سلولهاي آنتیپودي روي پوستامنت واقع شدهاند. پوستامنت برجسته میشود و آنتیپودها را به طرف مرکز کیسهرویانی می-راند.
یافتههاي نوین در علوم زیستی جلد 1، شمارة 70/70 Nova Biologica Reperta 1 (2): 67-76 (2015) 67-76 :2
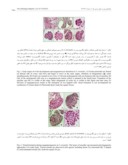
شکل -1 مراحل اولیه تکوین تخمک و تشکیل مگاسپوروسیت در (A .C.orientalis پریموردیومهاي تخمکی در سطح شکمی دیوارة تخمدان (Ov) تشکیل می-شوند و خمیدگی را از همان مراحل اولیه آغاز میکنند. بنیانگذاري پوسته (▲) نیز در همین زمان رخ میدهد؛ (B تخمکها در دو ردیف در داخل برچه مستقر هستند؛ (C چندین سلول آرکئوسپوري در رأس خورش وجود دارد؛ (D یکی از سلولهاي آرکئوسپور بهعنوان مگاسپوروسیت (mmc) عمل میکند، مگاسپوروسیت در لایه چهارم است. شکلگیري کلاهک خورش (NC) در این مرحله مشهود است. پوسته داخلی (ii) تخمک در این شکل و شکلهاي بعدي دیده میشود؛ (E با تقسیم سلول مگاسپوروسیت دو سلول دیاد هاپلوئید شکل میگیرد. در این شکل دیاد بدون سیتوکینز دیده میشود؛ (F دیاد خطی طولی؛ (G دیاد خطی عرضی. شاخص 20µm است.
Fig. 1. Early stages of ovule development and megasporocyte formation in C.orientalis. A) Ovular primordia are firmed on abaxial side of ovary wall (Ov) and begin to curve at the early stages. Initiation of integuments (▲) starts simultaneously; B) Ovules are located in two rows; C) Several archeosporial cells are formed at the top of nucellus; D) One of the archeoporial cells acts as a megasporocyte (mmc) which is situated in the fourth layer. Formation of the nucellar cap (NC) is visible at this stage. Inner integument of ovule (ii) is visible in this figure and later ones; E) Division of megasporocyte results in the formation of two haploid cells. Dyad formation is not accompanied by
cytokinesis; F) Linear dyad; G ) Horizontal dyad. Scale bar equals 20 µm.
شکل -2 تشکیل تتراد در طی مگاسپوروژنز در .C.orientalis لایههاي کلاهک خورشی افزایش یافته و پوستهها حدوداً به 3/4 جسم تخمک میرسند. انواع تتراد به شرح زیر در نمونه تحت بررسی دیده شد: تتراد خطی طولی((A، تتراد خطی عرضی (B)، تتراد T شکل (C) و تترادها از نوع تتراهدرال (D) مشاهده شد. شاخص 20µm است.
Fig. 2. Tetrad formation during megarporogenesis in C.orientalis. The layers of nucellar cap increased and integuments approaches 3/4 ovular body. Various tetrads are observed in this species including linear (A), horizontal (B), T-shaped
(C) and tetrahedral tetrads (D). Scale bar equals 20 µm.
یافتههاي نوین در علوم زیستی جلد 1، شمارة 67-76 :2
سلولهاي آنتیپود، بزرگترین سلولهاي کیسه رویانی هستند. سیتوپلاسم آنها متراکم است و هسته کروي یا بیضیشکل دارند. شمار این سلولها و تعداد هسته آنها طی تکوین افزایش نمییابد اما بهطورقابل ملاحظهاي بزرگ میشوند و داراي هسته حجیمی هستند که نمایانگر وجود پلیپلوئیدي در آنها است. یک یا دو عدد از سلولهاي آنتیپودي بعد از لقاح همچنان حضور دارند و درواقع پایا هستند. توسعه آنتیپودها همراه با واکوئلسازي در سیتوپلاسم و افزایش در ابعاد هسته و هستک است (شکلهاي .(3C-4E
هستههاي قطبی در کیسه رویانی در رأس سلولهاي آنتیپودي قرار گرفته (شکل (3E و بهزودي به هم ملحق میشوند تا هسته ثانویه و سلول مرکزي (شکل (4A را تشکیل دهند. سلول مرکزي "هسته ثانویه" ارتباطات سیتوپلاسمی با آنتیپودها برقرار میکند، در واقع این ارتباطات سیتوپلاسمی بین کلیه سلولهاي کیسهرویانی برقرار است. سپس سلول مرکزي مهاجرت میکند و در زیر دستگاه تخمزا قرار میگیرد. در C.orientalis هسته ثانویه قبل از لقاح تشکیل میشود. به دنبال ورود لوله گرده به تخمک (شکل (4B و انجام لقاح مضاعف، سلول تخم و سلول مادر آندوسپرم تشکیل میشود. زیگوت نسبت به سلول تخمزا متراکمتر است (شکل .(4C یک یا دو عدد از آنتیپودهاي پلیپلوئید بعد از لقاح تا مدتی پایا میمانند (شکلهاي 4E و (4Cو احتمالاً در مرحله اندوسپرم سلولی تحلیل میروند. با اولین تقسیم سلول تخم، سلول رأسی و قاعده-اي شکل میگیرد (شکل .(4E ادامه تقسیمات، رویان را ایجاد میکند (شکل .(4D با تشکیل کیسهرویانی و سلولهایی که قابلیت لقاح را دارند، یعنی تخمزا (هاپلوئید) و هستهدوتایی (دیپلوئید)، تکوین تخمک به پایان میرسد. طی بلوغ کیسه-رویانی کلاهک خورشی تحلیل میرود. در این گونه تکوین کیسهرویانی از تیپ پلیگونوم پیروي میکند. کیسه رویانی در طی بلوغ گلابیشکل شده و بسیار عریض میگردد (شکلهاي .(3C-4E در C.orientalis دستجات آوندي در خورش و پوستهها مشاهده شد (شکل .(3B
71/71 Nova Biologica Reperta 1 (2): 67-76 (2015)
بحث
در تیرة آلالهایان توالی بنیانگذاري مارپیچی اندامهاي گل یک الگو است (Ren et al ., 2009) و یک سطح سیستماتیکی پایین را نشان میدهد .(Endress et al., 1995) در بازال آنژیوسپرمها تخمکهاي پرخورش غالب هستند ( Ren et al., 2004) و تیرة آلاله در بین تاکسونهاي ابتدایی دولپهاي، حجیمترین خورش را دارد. وضعیت دوپوستهاي و میکروپیل آندوستومیک ویژگیهاي ابتدایی هستند و میکروپیل bistomic و تخمک تکپوستهاي ویژگیهاي اشتقاقیافته هستند .(Wang & Ren, 2007) در نمونه تحت مطالعه تخمک دوپوستهاي است و ابتدا پوسته درونی و سپس پوسته بیرونی بنیانگذاري میشود. بهعلاوه، پوسته بیرونی از رشد زیادي برخوردار است و تعداد لایههاي آن بیش از دو عدد است، درحالیکه پوسته درونی دولایه است. در پوستههاي چند-لایهاي، اپیدرم و پارانشیم دیده میشود (رضانژاد و چهرگانی،.(1387
در تشکیل میکروپیل فقط پوسته درونی دخالت دارد. بهعبارتی؛ میکروپیل اندوستومیک است که نوعی ویژگی ابتدایی محسوب میشود. در تیرة آلاله تقسیمات مماسی سلولهاي اپیدرمی به تشکیل کلاهک خورشی منجر میشود. درجه نمو کلاهک، یک ویژگی مخصوص گونه است. کلاهک خورشی در C.orientalis دو تا چهار لایه است. در خانوادة آلاله کلاهک خورشی میتواند موجب تشکیل اپیستاز شود که بافتی با دیوارة سلولی ضخیم است و در دانههاي نیز بالغ پایدار میماند (Ren .et al., 2009) ساختارهاي هیپوستاز و پوستامنت در گونه C.orientalis مشاهده شد. درواقع در تخمکهاي پرخورش، سلولهاي بخش نزدیک (به سلولهاي آرکئوسپوري) به پوستامنت، سلولهاي ناحیه دور به دسته آوندي و سلولهاي ناحیه میانی (منطقه مرکزي) به پودیوم و هیپوستاز نمو مییابند. همه این ساختارها درحالیکه عملکردهاي مشابهی انجام می-دهند ازنظر الگوي تشکیل متفاوت هستند. پوستامنت بافتی با خاستگاه خورشی است که به صورت یک ستون از سلولهاي طویل مستقر در بخش زیرین انتهاي آنتیپودي کیسهرویانی قرار گرفته است. سلولهاي پوستامنت از سلولهاي اطراف به-
یافتههاي نوین در علوم زیستی جلد 1، شمارة 67-76 :2
وسیله سیتوپلاسم متراکمتر و دیوارههاي رنگ پذیرتر قابل-تشخیص هستند. پوستامنت در تیرة آلاله از لایههاي محدودي تشکیل شده است و به سمت هیپوستاز توسعه یافتهاست و از نظر ساختاري با سلولهاي پروکامبیومی قابل مقایسه است (Joshi
& Venkateswarlu, 1935; Mück, 1980; Kamelina & Proskurina, 1987a,b; Alimova, 1990; Shamrov .& Nikiticheva, 1992)
وجود چندین آرکئوسپو، از ویژگیهاي تیرة آلاله و به طور کلی از خصوصیات تخمکهاي با خورش حجیم و چندلایهاي است. اما فقط یکی از آنها به سلول مادر مگاسپور (mmc) تمایز می-یابد. مگاسپوروژنز با الگوي رایج در نهاندانگان دولپه انجام میشود. مطالعه روند مگاسپورانژ نشان میدهد که شکل تترادها در این تیره تنوع بسیار زیادي دارد که با گزارشهاي پیشین
همسو است (Kozieradzka-Kiszkurno & Bohdanwicz,
.2003)
در C.orientalis مگاسپور پس از تشکیل به کیسهرویانی نمو می کند و الگوي تکوین کیسه رویانی از تیپ پلیگونوم پیروي می کند که تیپ رایج در اکثر نهاندانگان است (Kordyum, .1959) انواع دیگر تکوین کیسهرویانی در این تیره گزارش شده است. Adonis تکوین از نوع Allium را نشان داده است
.(Uluce & Ünal, 1962; Bhandari, 1996) در Trollius
acaulis Lindl. تیپ دو اسپوري و در T. europeus تیپ مونواسپوري گزارش شده است (Bhandari & Kaplin,
1964)؛ درحالیکه در Thalictrum angustifolium L. var. flavum کیسه رویانی تتراسپوري 16 هستهاي منطبق بر تیپ ethrumPyr گزارش شده است (Picci, 1964)، اما در
T. javanicum Grun. تکوین گامتوفیت ماده از نوع مونواسپوري 8 هستهاي و تیپ پلیگونوم است که با گزارش ما دربارة C.orientalis همسویی دارد.
براساس نتایج، در C.orientalis سلولهاي آنتیپود در رأس پوستامنت قرار میگیرند که با گزارشهاي در C.regalis L. همسویی دارد .(Unal & Vardar, 2006) در دو گونه اخیر همانند دیگر تخمکهاي پرخورش، سلولهاي ناحیه پروکسیمال به پوستامنت نمو مییابد که نشان ویژة ساختاري تیرة آلاله است .(Batygina & Shamrov, 1999) پوستامنت رشد میکند و
72/72 Nova Biologica Reperta 1 (2): 67-76 (2015)
سلولهاي آنتیپود را به مرکز کیسه رویانی میراند و ورود لوله گرده درحالی صورت میگیرد که آنتیپودها کاملاً به وسط کیسه رویانی پیشروي کردهاند. طول عمر آنتیپودها در گیاهان مختلف متفاوت است و با تکوین عملکرد ساختارهاي خورشی دیگر در ارتباط است. در بین سلولهاي کیسهرویانی میزان پایداري آنتیپودها بیشترین تنوع را نشان میدهد. به طور بسیار رایج آنها مدت کوتاهی پساز لقاح تحلیل میروند، اما می-توانند تا مدت طولانی حتی بعد از لقاح نیز باقی بمانند
.(Hasitschka- Jenschke, 1962; Ünal et al., 1997) در
این خانواده با توجه به اندازهگیري ابعاد هسته وجود سلولهاي آنتیپود و وجود تیپهاي متنوعی از پلیپلوئیدي در سلولهاي آنتیپود گزارش شده است. پدیدة پلیپلوئیدي ممکن است در-نتیجه اختلال در میتوز باشد؛ براي مثال در برخی از گونههاي
nomenA (Hasitscheka-Jenschke, 1959; Trela,
1963; Ünal, 1978) یا حاصل از اندومیتوز باشد،مثلاً در (Turala-Szybowska, 1980) Ranunculus peltatus و
.(Tschermak-Woess, 1956) Aconitum
