







بخشی از مقاله
بررسی اپیدرمی و اکولوژی ویروس ها
مقدمه
به منظور تبدیل اطلاعات گلخانه ای و آزمایشگاهی موجود در مورد ویروسها به یک تصویر کلی از طبیعت آنها یعنی اکولوژی ( از وازه یونانی Oikos یعنی خانه وLogos به معنی سخنرانی )، که همیشه متغیر است نیاز داریم.
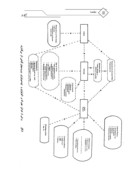
ویروسهای گیاهی برای بقاء باید دارای : (1) یک یا چند گونه گیاهی میزبان برای افزایش؛ (2) روشی مؤثر برای انتشار و ایجاد آلودگی در گیاهان میزبان جدید و ( 3) ذخیره ای از گیاهان میزبان مناسب سالم به منظور انتشار بیماری باشند.موقعیت واقعی هر ویروس معین در یک محل خاص یا در مقیاس جهانی، نتیجه برهمکنشهای پیچیده بین بسیاری از عوامل فیزیکی و بیولوژیکی خواهد بود(شکل1، پائین صفحه) درک و شناخت اکولوژی یک ویروس در یک محصول و محل معین، جهت
توسعه روشهای مناسب برای کنترل بیماری ویروسی لازم و ضروری است. همانند اغلب پارازیتهای اجباری، عوامل اکولوژیکی عمده ای که باید در نظر گرفته شوند، بیشتر شامل همان روشهایی است که موجب انتشار ویروس از گیاهی به گیاه دیگر شده و نیز راههایی است که سایر عوامل روی انتشار ویروس تأثیر میگذارند.
بررسی نموداری، در وهله اول، ارتباط عواملی که توصیف خواهد شد را در یک چشم انداز اکولوژیکی، با اشاره به پیچیدگی تعاملهای آنها، نشان خواهد داد. آنگاه مداخله و تعاملهای کیفی عوامل اکولوژیکی را میتوان بررسی نمود. سپس با خلاصه ای در مورد چگونگی گسترش کمی آلودگی در گیاهان زراعی یامورد حمله قرار گرفتن آنها یا جمعیتهای گیاهی دیگر دنبال خواهد شد. این، نکته اصلی اکولوژی کمی است که اپیدمیولوژی نامیده میشود(ازکلمه یونانی Epi به معنی"برروی"و Demos برای"مردم وجمعیت"وLogosبه معنی سخنرانی ).اپیدمیولوژی از کمیت و مدل سازی صحبت میکند. و هم اکنون به طور چشمگیر مورد حمایت فنون جدید ردیابی در گیاهان و ناقلین ، قرار دارد.
نمودارها
بیماری ویروسی در یکایک گیاهان و در جمعیتهای گیاهی در نتیجه عکس العمل بین ویروس، میزبان و محیط ناشی میشود. این موضوع به صورت مثلث بیماری در شکل1 به سادگی نشان داده شده است (Hull,1991b ).با وجود این، این چنین تعاملها و اثر محیط بسیار پیچیده تر هستند.
(شکل 2) شامل گروههای اصلی عوامل مداخله کننده در اکولوژی ویروسها میباشد که نمایانگر اکوسیستم ویروسی است. عوامل اصلی ویروسها هستند، که تمایل دارند در همه جا ظاهر شوند و اگر موجب وقوع خسارت بشوند باید تحت کنترل قرار گیرند. گیاهان میزبان به محصولات هدف که باید حفاظت شوند و منابع آلودگی که ویروسهای گیاهی اکثراً از آنها نشأت میگیرند، اشاره میکند.
اکولوژی ویروسها
اثری که ویروسها میتوانند در گیاهان به جای بگذارند با مشخصات زیست شناسی آنها، معمولاً روابط ناقلی و اختصاصی بودن میزبان، تعیین میشود. ویروسها و حتی نژادهای ویروسی ممکن است از نظر تهاجم و بیماریزایی تفاوت فاحشی داشته باشند و حتی ویروسهایی که باعث بیماریهای مشابه میشوند از نظر اکولوژی ممکن است کاملاً متفاوت باشند. بنابراین تشخیص دقیق نیازمند تحقیق روی اکولوژی و اپیدمیولوژی و کنترل نهایی ویروس میباشد. ویروسها به طور فزاینده در همه جا در هر گونه گیاهی از قارچ، جلبکها و سروها گرفته تا تمامی انواع گیاهان دانه دار کشف یا ظاهر میشوند.
اطلاعات روزافزون از تحقیقات مداوم انجام گرفته روی ویروس، از روشهای ردیابی پیشرفته و بررسی مداوم وجود ویروسها در گیاهان زراعی و پوشش گیاهی وحشی حاصل میشود. معلوم میشود که تک تک ویروسها دارای طیفهای میزبانی وسیعی بوده و پراکنش وسیع تراز آنچه که برای مدتها تصور میشد، دارند.برای نمونه، چنین بررسیهای انجام گرفته در حبوبات در دو دهه اخیر در سراسر آسیای غربی و آفریقای شمالی توسط ایکاردا ( مرکز بین المللی تحقیقات کشاورزی در نواحی خشک ( ICARDA ) در حلب سوریه به وجود آمدن حداقل 12 نوع ویروس ر
ا در این گیاهان مشخص کرده اند و معلوم شده است که اکثر این ویروسها دارای پراکندگی وسیعی بوده و اغلب از احتمال وقوع بالایی برخوردار میباشند. برخی از این ویروسها قبلاً گسترش یافته اند و اکثر آنها، به علت سرایت از طریق بذر، خطری نسبت به برنامههای به نژادی
زراعی محسوب میشوند. به عنوان نمونه، بروموویروس پیسک باقلا که با سوسک منتقل میشود ابتدا به صورت تصادفی در انگلستان در سال 1951 ردیابی شد و مدتها تصور میشد که تنها از نظر علمی و آکادمیک مهم است. هم اکنون معلوم شده است که در مراکش، الجزایر، تونس، سودان، مصر، و سوریه نیز یافت میشود. این ویروس در مراکش در 56% مزارع بازرسی شده باقلا، با حداکثر وقوع 33% در یک مزرعه، بوجود میآید. این ویروس نخود ایرانی، عدس و نخود
فرنگی را نیز آلوده میکند. نانوویروس زردی نکروتیک باقلا، که ابتدا در اوائل دهه 1990 از سوریه گزارش گردید، بلافاصله معلوم شد که سبب اپیدمی ناگهانی ویروسی در مصر در سال 1992 بوده است. مسترویروس کوتولگی کلروتیک نخود ایرانی که در سال 1993 در هند گزارش گردید، اخیراً به عنوان عامل یک ناهنجاری مهم در باقلا، که مشابه برگ قاشقی لوبیا بوده و برای مدتهای طولانی در سودان شناخته شده است، ردیابی شد. این ویروس هم اکنون در یمن و برخی کشورهای دیگر یافت میشود.
چند نمونه از چگونگی افزایش سریع شیوع ویروسها و اهمیت یا افزایش پراکندگی آنها از منطقه پیدایش وجود دارند: (1) به وسیله انسان از طریق عملیات زراعی (2) از راه تجارت روزافزون بین مناطق و قارهها و (3) بین المللی شدن فزاینده به نژادی گیاهان زراعی از طریق مبادله در مقیاس وسیع (الف) ژرم پلاسم و مواد اصلاحی برای انجام آزمایش چندمکانی و (ب) ازدیاد تجارتی مواد تکثیر گیاهی. حتی برخی از ویروسها ممکن است، بدون دخالت انسان، در مسافتهای طولانی، به عنوان مثال به همراه حشرات، از طریق جریانات هوا و باد و پخش بذر توسط آب، باد و حیوانات گسترش یابند. کلیه ویروسها، با توجه به مفهوم تئوری " اجتناب ناپذیری استقرار" بالاخره ممکن است به هر جائی راه یابند اما ظهور آنها فقط در شرایط اکولوژیکی مساعد، امکان پذیراست. بنابراین، در کشاورزی امروزی ویروسها از اهمیت روزافزونی برخوردار میباشند. پیدایش بیماریهای ویروسی جدید ممکن است در نتیجه تغییرپذیری ویروس ناشی شود. ویروسها از نظر دامنه میزبانی، قدرت بیماریزایی و داشتن ناقل اختصاصی، همیشه در حال تغییر هستند. ویروسها به طرق مختلف به شرح ذیل تغییرمی یابند: (1) جهش و گزینش از راه سازگاری مانند گذر از میزبانهای ویژه، انواع جدید گیاه یا کولتیوار محصول، (2) تعامل مستقیم نژادهای ویروس یا
ویروسهای مختلف در آلودگیهای مخلوط که منجربه (الف) کپسیدی شدن ناهمگن، نوترکیبی کاذب یا حتی(ب) نوترکیبی ژنی بین ویروسهای مختلف میشوند.
اطلاعات اندکی در مورد اثر مستقیم شرایط اقلیمی بر روی ویروسها وجود دارد. برخی از ویروسها ممکن است در دمای بالا فعال باشند، در صورتی که ویروسهای دیگر دماهای پائین تر را ترجیح
میدهند یا در دماهای پائین، بیماری زاتر میشوند. در دمای بالا، ویروسها یا نژادهای مقاوم به گرما، مانند کوکوموویروس موزاییک خیار ( CMV )، ممکن است غالب شوند. اکثر ویروئیدها فوق العاده بیماریزا هستند، به غلظتهای فوق العاده بالا میرسند و علائم را در دماهای بالاتر از 30 درجه سانتی گراد، در مقایسه با دماهای پائین تر، زودتر بروز میدهند. با وجود این، اثر دما اغل
ب بر روی میزبان، و نه ویروس، است. مثالی در این مورد، تغییر علائم از واکنش لکههای موضعی نکروتیک در 20 درجه سانتی گراد در گیاه Nicotiana glutinosa مایه زنی شده با ویروس موزاییک توتون، به موزاییک سیستمیک در 30 درجه است. چند روز پس از پائین آوردن دما به 20 درجه، موزاییک به نکروز سیستمیک تغییر مییابد.
عوامل بیولوژیکی
1)خصوصیات ویروسها و گیاهان میزبان آنها
1-1 پایداری فیزیکی و غلظت نهایی ویروسها
در ویروسهای وابسته به انتقال مکانیکی، احتمال بقاء و انتشار ویروسی که داخل و خارج از گیاه پایدار بوده و غلظت آن نیز در بافت خیلی بالاست، بسیار بیشتر از ویروسی است که پایداری و غلظت کمتری دارد. به نظر میرسد که بقا و انتشار برخی ویروسها، عمدتاً به پایداری زیاد و مقادیر بالای تولید شده آنها در بافتهای آلوده بستگی دارد. به عنوان مثال، TMV ممکن است برای مدت طولانی در مواد گیاهی مرده و در خاک دوام پیدا کند. به این ترتیب منبعی برای آلودگی محصولات بعدی فراهم میگردد.TMV همچنین در فرایند تولید توتون از بین نرفته و در سیگارها نیز یافت میشود.
1-2 میزان حرکت و انتشار در گیاهان میزبان
ویروسها یا نژادهای ویروسی که در گیاه میزبان و از محل آلودگی به کندی حرکت
می کنند، در مقایسه با ویروسهایی که دارای سرعت حرکت بیشتری هستند، از قدرت انتشار و بقای کمتری برخوردار میباشند.سرعت حرکت ویروس در گیاه مهم بوده و نسبت به طول عمرهر یک از گیاهان میزبان اندازه گیری میشود. ویروسهایی که درختچهها و یا درختان ( با عمر طولانی ) را آلوده میکنند، در مقایسه با ویروسهایی که در گیاهان یکساله آلودگی ایجاد میکنند،
میتوانند با سرعت بسیار بسیار کمتری در میزبان خود حرکت کنند. همچنین ویروسهایی که قادرند به درون بذر حرکت کرده و در آنجا بقا یابند، در انتشار و بقای خود دارای مزیت مهمی میباشند.
1-3 شدت بیماری
احتمالاً ویروسی که در میزبان خود به سرعت توسعه پیدا کرده و بیماری سیستمیک تولید نماید به طوری که موجب مرگ میزبان خود گردد، در مقایسه با ویروسهایی که بیماری ملایم یا متوسطی ایجاد میکنند، از احتمال بقای بسیار کمتری برخوردار میباشد. در بیماری ملایم و یا متوسط ، میزبان میتواند بقاء یافته و به صورت مؤثرتری تولید مثل کند. به نظر میرسد در مزرعه علیه نژادهایی که موجب مرگ سریع میزبان خود میگردند، انتخاب طبیعی وجود دارد.
1-4 تغییرپذیری و انتخاب نژاد
احتمال تولید نژادهای جدیدویروس بر اثر موتاسیون و یا نوترکیبی حقیقی و کاذب
اسید نوکلئیک در ویروسهایی که ژنوم چند بخشی دارند، وجود دارد. این نژادهای جدید ممکن است به طور ناگهانی ظاهر شده و خسارات شدیدی به محصولی که تا به آن زمان آن ویروس خاص راتحمل میکردند و یا آلوده نمی شدند وارد نمایند.
مثلاً ویروس کوتولگی ذرت سالها در نیشکر شیوع داشت، و خسارات زیادی را در بسیاری از کشورها موجب شد تا اینکه ابتدا ارقام متحمل و سپس مقاوم به این ویروس معرفی گردید. نژادهای متعددی از این ویروس وجود داشت ولی به ندرت ذرت را آلوده میکردند مگر اینکه ذرت در مجاورت نیشکر کشت میشد. اخیراً شرایط در بسیاری از نقاط ایالات متحده به شدت تغییر کرده است وویروس ذرت را آلوده میکند و خسارات زیادی در بسیاری از نواحی که کاملاً از مزارع نیشکر جداهستند ایجاد کرده است. تغییر در شدت بیماری بر اثر افزایش ایجاد نژادهای جدید ویروس است.
به طور کلی میزان توانایی جهش یک ویروس برای تولید نژادهای جدیدی که بتواند به صورت مؤثری با تغییرات محیطی سازگار باشند، ممکن است تأثیر زیادی بر بقاءو انتشار ویروس مورد نظر داشته باشد. مشخص شده است که عوامل زیر تحت شرایط خاصی، در فشارهای انتخابی نقش مهمی ایفا مینمایند: (1) گونه و رقم گیاه میزبان؛ (2)گونه و نژاد ناقلین بی مهره؛ (3) تغییرات فصلی و آب و هوایی، به خصوص تغییر در درجه حرارت. در فصول مختلف ممکن است نژادهای غالب یک
ویروس در یک محصول متفاوت باشد و (4) عملیات کشاورزی، به ویژه در محصولاتی که به صورت رویشی ازدیاد پیدا میکنند، میتواند موجب انتخاب نزادهای خاصی از یک ویروس شود.
1-5 دامنه میزبانی
ویروسها از نظر دامنه میزبانی گونههای گیاهی که میتوانند آلوده کنند، بسیار متفاوت هستند. به نظر میرسد بعضی از ویروسهای عامل بیماری در توت فرنگی، محدود به جنس Fragaria باشند. در حالیکه ویروسهای دیگر ممکن است بتوانند طیف وسیعی از گیاهان را آلوده نمایند. به عنوان مثال ویروس موزاییک خیار(CMV ) میتواند گونههای متعلق به بیش از 100 خانواده گیاهی را آلوده نماید. به نظر میرسد عامل بقاء در ویروسهایی که دارای دامنه میزبانی محدودی هستند، ناشی از داشتن میزبانهای چندساله و ازدیاد رویشی آنها یا انتقال مؤثر ویروس از طریق بذر باشد.
تنوع و گوناگونی میزبانهای یک ویروس، فرصت زیادتری برای حفظ، بقا و انتشار بیشترویروس فراهم میآورد. ویروسهایی که دارای میزبانهای زینتی چند ساله مثل گل کوکب و سوسن هستند، همانند گونههای زراعی و باغی در سراسر دنیا گسترش یافته اند. علفهای هرز، گیاهان وحشی، بوتههای پرچین، درختان زینتی و درختچهها ممکن است به عنوان منبع نگهدارنده ویروس عمل کنند. اهمیت واقعی انواع مختلف میزبانهای ویروسی برای محصولاتی که در مجاورت آنها کاشته میشوند، به عوامل متعددی به ویژه حضور ناقلین بی مهره فعال بستگی دارد.
2)انتشار
انتشار ویروسها توسط ناقلین هوازاد یا ساکن خاک، یا بوسیله بذر و گرده و یا در مسافتهای طولانی و در نتیجه فعالیتهای انسان، نقش کلیدی و بسیار مهمی در اکولوژی ویروسها دارد.
مقدار و دامنه انتشار آلودگی از هر کانون آلودگی به شیب آلودگی معروف است. همان گونه که انتظار میرود، تعداد گیاهان آلوده با دور شدن از کانون آلودگی کاهش مییابد. تندی یا ملایمی شیب آلودگی بستگی به نوع ناقل، نحوه انتقال ویروس و عواملی که حرکت و فراوانی ناقل را کنترل میکنند دارد. شیب ملایم شامل انتشار در فواصل قابل ملاحظه ای است که توسط ناقلین با تحرک زیاد ایجاد میشود، در حالی که شیب تند که انتشار در فواصل کوتاهتری است توسط ناقلین با تحرک کمتر یا انتقال بوسیله تماس ریشه یا برگهای مجاور بوجود میآید. ناقلین بالدار مثل
شتهها در جهت باد خیلی آسانتر منتقل میشوند تا در جهت مخالف باد، به همین دلیل شیب آلودگی در جهت باد معمولاً ملایم ولی در جهت مخالف باد معمولاً تندتر است.
ماهیت انتقال ویروس به طرق پایا و ناپایا به وسیله ناقل نیز در نحوه انتشار ویروس مؤثر است. هنگامیکه ناقلی یک ویروس پایا را انتقال میدهد، ناقل برای مدتی طولانی و در فاصله دوری آلوده به ویروس خواهد بود، در حالی که در مورد ویروس ناپایا، ناقل برای زمان و فاصله کوتاهی ویروس دار است.
2-1 ناقلین هوازاد
حشرات ناقلی که به صورت مکشی از عصاره گیاه تغذیه کرده و میتوانند پرواز کنند و به خصوص شتهها، مهم ترین عوامل انتشار و بقای ویروسهای گیاهی میباشند. شکل و نحوه انتشار ویروس در محصول مورد نظر و میزان گسترش آن ، به عوامل بسیاری بستگی خواهد داشت که عبارتند از: (1) منبع مایه. منشأ مایه میتواند از بیرون محصول، تک بوتههای بیمار داخل مزرعه و یا از بقایای محصول قبلی باشد؛ (2) میزان پتانسیل مایه موجود؛ (3) طبیعت و رفتار ناقل؛ برای مثال در شتهها، این که آیا افراد بالدار محصول را کلونیزه میکنند یا خیر؛ (4) آیا ویروس در ناقل خود به صورت ناپایا، نیمه پایا یا پایا است؛ (5) زمان فعال شدن ناقلین در مقایسه با دوره زندگی محصول و (6) شرایط آب و هوایی.
در صورت عدم انتقال با بذر،آلودگیهای پراکنده در مزرعه احتمالاً از یک منبع خارج از مزرعه تأمین شده و توسط ناقلینی که پرواز میکنند، منتقل میگردند. تجمع گیاهان واگرفته به صورت گروهی، نشان دهنده گسترش آلودگی از منایع داخل محصول است. به ویژه شتههای بالدار عبوری که در مجموع محصول را کلونیزه نکرده و از گیاهی به گیاه دیگر حرکت مینمایند،می توانند در گسترش ویروسهای انتقالی به روش ناپایا اهمیت داشته باشند. هرچند این شتهها ممکن است نسبت کوچکی از کل جمعیت را تشکیل دهند، ولی میتوانند ویروسها را از بیرون مزرعه یا از گیاهان واگرفته موجود در مزرعه کسب کرده و ضمن حرکت برای یافتن گیاهان مناسب، موجب انتشار و گسترش سریع ویروس گردند. شتههای کلونیزه کننده نیز در صورت حرکت بین محصول، مهم خواهند بود.
در مناطق معتدله جمعیت شتهها روی گیاهان زراعی ممکن است به مدت چندین نسل به صورت پارتنوژنیک (غیرجنسی) سریعاً افزایش یابند. شفیرههای شته سبز هلو از چهار مرحله یا پوست اندازی میگذرند و شتههای بزرگسال، مادههای زنده زا میباشند.اکثر این مادهها شتههای بی بال هستند که کلونیها را بوجود میآورند. اگر تراکم این شتهها بیش از اندازه زیاد باشد ( ازدحام ) بزرگسالان بی بال بوجود میآیند.اینها برای جستجوی منابع غذائی جدید ممکن است به حرکت درآیند و از اهمیت ویژه ای برای پخش ویروسها برخوردار هستند. همچنین، شتههای بالدار در
شرایط آب و هوایی خنک، در پائیز که میزبانهای علفی از بین میروند و میزبانهای جایگزین برای زمستان گذرانی باید مورد جستجو قرار گیرند، تولید میشوند.میزبانهای زمستانی عمدتاً درختچهها و درختهای وحشی، مانند شمشاد و بداغ جنگلی برای زمستان گذرانی شته Aphis fabae ،گونههای بید برای شته Cavariella aegpodii و گونههای آلو و دیو خار برای شته سبز هلو هستند. شتههای ماده بی بال روی این میزبانها تولید میشوند، با نرهای بالدار جفت گیری میکنند و برای تکمیل چرخه زندگی خود تخم میگذراند. در بهار هنگامی که برگهای جوان
رشد میکنند، تخمها باز میشوند تا تشکیل مادههای بالدار، به نام فونداتریکس، را بدهند. مهاجران بالدار پس از یک یا دو نسل روی میزبانهای زمستانی، در اواخر بهار یا اوایل تابستان تولید میشوند و اینها به میزبانهای تابستانی کوچ میکنند. به هنگام زمستانهای ملایم ، شتهها ممکن است به میزبانهای زمستان گذران حرکت نکنند اما به توسعه خود به طریق زنده زائی روی میزبانهای تابستانی ادامه میدهند. چنین توسعه غیرچرخه ای در مناطق گرمسیری معمول میباشد.
مگسهای سفید مانند مگس سفید سیب زمینی شیرین گرمسیری یا پنبه و مگس سفید گلخانه دارای چهار مرحله لاروی هستند که فاقد تحرک بوده و از آوند آبکش زیر برگها تغذیه میکنند. مرحله چهارم لاروی به پوپاریوم ( آخرین پوسته لاروی سخت شده ای که شفیره را میپوشاند) تبدیل میشود که بزرگسالان پرواز کننده از آنها بیرون میآیند. تخمها به صورت الگوی شبه هلالی در حالی که حشره ماده شکم خود را به هنگام تغذیه با برگ تماس میدهد، گذاشته میشوند. جمعیت این مگسها ممکن است به سرعت به نسبتهای اپیدمیکی برسد.
رشد جمعیت ناقلین و انتشار ویروس بیشتر به فیزیولوژی میزبان و همین طور به نوع گیاه محصول، تراکم گیاه، سن و تغذیه ناقلین بستگی دارد. آلودگی ویروسی نیز فیزیولوژی میزبان و به همین ترتیب قابل تغذیه بودن آن را برای ناقل تحت تأثیر قرار میدهد و تغییر در رنگ میزبان نیز ممکن است دارای نقشی باشد. آلودگی مویز سیاه با عامل تغییر رنگ (نپوویروس تغییررنگ مویز)، به طور چشمگیری آلودگی بیشهها توسط ناقل آن ویروس، کنه گالی Cecidophyopsis(Phytoptus) ribis را افزایش میدهد.
رفتار بندپایان تحت تأثیر دما، نور و سرعت باد قرار میگیرد. شکل 3 رفتار پروازی شتهها را نشان میدهد. به هنگام پرواز، شتههای بالدار ممکن است مکرراً فرود آمده و تغذیه کرده و مجدداً برای یافتن میزبانهای مناسب چندین پرواز کوتاه دیگر معمولاً 1 تا 2 متری انجام دهند. هنگامیکه افراد شته سبز هلو چنین پروازهائی انجام میدهند ممکن است روی 7 تا 10 گیاه مختلف لارو بگذارند. این چنین حرکات شتهها را برای پخش ویروسهای ناپایا در مسافتهای کوتاه حتی در گیاهان و محصولاتی که میزبانهای شته نیستند، ایده آل میکند. بین شتههای ویزیتور مهاجر (کلونیزه نکننده )، شتههای غیرمهاجر و ساکن ( کلونیزه نکننده) و شتههای مهاجر ( کلونیزه کننده) باید تفاوتی قائل شد. شتههای بی بال میتوانند، زمانی که از گیاهی به گیاه دیگر میخزند، ویروسها را انتقال دهند اما شتههای بالدار ویروس را به مسافتهای بلند ممکن است حمل کنند.
شکل 3 _ نمایش شماتیک رفتار پروازی شتههای بالدار( Moerick,1955 )
.
این حقیقت که اکثر مطالعات اپیدمولوژیکی روی ویروسهای شته زاد انجام گرفته است اهمیت این گروه از ناقلین را نشان میدهد.
2-2 ویروسهای خاکزاد
ویروسهایی که ناقل آنها شناخته نشده است
TMV یکی از معدودترین ویروسهایی است که بدون کمک هر نوع ناقل شناخته شده، به میزبان زیادی در خاک انتقال پیدا میکند. پایداری این ویروس موجب دوام آن در بقایای گیاهی از فصلی
ته شده ای تدارند، در خاکهای اکوسیستم جنگلی آلمان گسترش یافته اند.
ویروسهای با ناقلین قارچی
مفاهیم اکولوژیکی انتقال با ناقلین قارچی، به روش حمل ویروس توسط قارچ بستگی دارد. ویروسهایی مثل ویروس موزاییک خاکزاد گندم، داخلهاگهای استراحتی
( پلاسمودیوفورومیستهای ناقل ) حمل میشوند. در اینهاگها ویروس میتواند به مدت چندین سال در خاکی که با هوا خشک شده باشد، دوام پیدا کند. در ویروسهایی که با قارچ انتقال مییابند، هر چند ممکن است ایجاد آلودگی چندین فصل طول بکشد، ولی چنان چه فقط یکبار مستقر شوند، ممکن است حتی در غیاب میزبان گیاهی مناسب برای مدت طولانی دوام بیاورند. انتشار موضعی این ویروسها توسط زئوسپورها وهاگهای استراحتی و از طریق آبهای موجود در خاک، مهم ترین عوامل مؤثر در حرکت آنها میباشند. ویروسهایی که با این روش انتقال پیدا میکنند، ترجیحاً دامنه میزبانی محدودی دارند.
از طرفی ویروسهایی که توسط قارچ Olpidium brassicae منتقل میشوند، مانند ویروس بافت مردگی توتون ( TNV) وویروس وابسته TNV ( STNV )، در سطحهاگها حمل میشوند. زئوسپورهایی که ویروس را حمل میکنند، احتمالاً فقط چند ساعت دوام میآورند. هرچند ممکن است ویروسهای آزاد در خاک ، توسط زئوسپورهایی که تازه رها شده اند، برداشته شده و انتقال پیدا کنند. ویروسهایی که توسط Olpidium brassicae انتقال مییابند، در خاک خشک شده با هوا دوام و بقا پیدا نمی کنند. به طور کلی این نوع ویروسها دامنه میزبانی وسیعی دارند و احتمالاً با انتقال مکرر به میزبانهای متوالی، در خاک دوام مییابند. ریزش آب، حرکت خاک و قطعات ریشه، احتمالاً در گسترش این ویروسها در خاک از محلی به محل دیگر نقش مهمی دارند. ثابت شده است که واریکوساویروس کوتولگی توتون در اسپورهای استراحتی به مدت بیشتر از 20 سال باقی میماند. این اسپورها در رابطه با واریکوسا ویروس رگبرگ درشتی کاهو و قارچ Polymyxa betae با بنی ویروس نکروز رگبرگ زرد چغندر ( BNYVV) در آب زهکشی شده یافت شده اند.
همچنین ویروسهای گیاهی که انتقال آنها با قارچ امکان پذیر است باعث نگرانی فزاینده در کشت عاری از خاک ( هیدروپونیک ) در محلولهای غذایی، بویژه زمانی که این محلولها چرخنده باشند، میشوند.
ویروسهای با ناقلین نماتدی
بین اکولوژی ویروسهایی که توسط نماتدها انتقال مییابند مانند توبراویروسها و نپوویروسها و ویروسهای با ناقلین هوازاد، تفاوت قابل ملاحظه ای وجود دارد. نماتدها با عمر طولانی و امکان داشتن دامنه میزبانی وسیع، قادرند در شرایط نامساعد و در غیاب گیاهان میزبان، برای مدت زمان قابل توجهی بقا یابند. نماتدهای ناقل، مرحله استراحتی مقاوم ندارند ولی با حرکت از میان پروفیل خاک ، میتوانند تحت شرایط نامناسب خاک دوام پیدا کنند. هنگامیکه خاک در تابستان گرم یا در زمستان سرد میشود، نماتدها به سمت نواحی زیرین خاک حرکت میکنند و با مناسب شدن شرایط برای آنها، به سطح خاک باز میگردند. بعضی از ویروسها مانند ویروس موزاییک آرابیس،
میتوانند طی زمستان در نماتد ناقل خود بقا یابند. حرکت اندک نماتد در خاک انتشار ویروسهای نماتدزاد را به 60- 30 سانتی متر در سال محدود میکند، و به وجود آمدن این ویروسها در مزرعه نیز معمولاً به صورت لکه ای است برای نمونه، حرکت نماتد به ساختمان منفذی، محتوای آبی و حرارت خاک بستگی دارد.
ویروسهایی که توسط نماتدها منتقل میشوند، معمولاً دردو خصوصیت مشترک هستند: 1) دارای دامنه میزبانی وسیعی به خصوص در میان گونههای علف هرز میباشند.2) در بسیاری از میزبانهای خود از طریق بذر نیز انتقال مییابند. بنابراین نماتدهای ناقلی که طی زمستان آلودگی خود را از دست بدهند، احتمالاً با جوانه زنی علف هرز آلوده شده در بهار، مجدداً ویروس را کسب مینمایند.
انتشار نماتدها در خاکهای دست نخورده، ممکن است کندترباشد. عملیات کشاورزی و احتمالاً آب زهکشی یا طغیان آب، میزان پراکنش این ناقلین را افزایش میدهد. همچنین کفش کارگران و آلایش سطحی ماشینهای کشاورزی با خاک آلوده، ممکن است موجب انتقال و جابه جایی نماتدها و در نتیجه ویروسهای همراه با آنها در فواصل کوتاه یا طولانی گردد. در اغلب موارد، الگوی ظهور و گسترش آلودگی در یک محصول، میتواند تا حدود زیادی به وضیعت ویروس و ناقل در خاک قبل از مرحله کشت بستگی داشته باشد. پس از کشت گیاهان دو یا چند ساله در مزرعه آلوده به نماتد، ظاهر شدن نشانههای الگوی آلودگی میتواند یک یا دو سال طول بکشد. زیرا نشانههای آلودگی برگی ممکن است یکسال بعد از وقوع آلودگی ظاهر شوند.گسترش موضعی بعدی می تواند موجب توسعه تدریجی قطعات آلوده شده در مزرعه گردد.
2-3 انتقال با بذر و گرده
انتقال با بذر و گرده میتواند اهمیت قابل توجهی در اکولوژی برخی ویروسها داشته باشد. بقا در بذر ممکن است برای ویروسهایی که فقط دارای میزبانهای یکساله هستند و نیز ویروسهایی که ناقلین بی مهره آنها ( مثل نماتدها) عموماً به کندی حرکت میکنند، از اهمیت ویژه ای برخوردار باشد. بعضی از ویروسها میتوانند در بذور دفن شده، حداقل یکسال دوام آورند.انتشار طبیعی بذور آلوده شده توسط باد و یا آب نیز ممکن است عامل مهمی در انتقال ویروس باشد.
2-4 انتقال بوسیله مهره داران
مهره داران، ویروسهای فوق العاده مسری را ممکن است روی خز یا پرهای خود حمل کنند. مثلاً سگها و خرگوشها میتوانند پوتکس ویروس ایکس سیب زمینی (PVX ) و پرندگان گاهی توبموویروس موزائیک توتون را حمل کنند. جانوران، بویژه پرندگان ممکن است عامل انتشار بذور آلوده به ویروس باشند. بذر ممکن است به مدت طولانی در دستگاههاضمه پرندگان مانده و به صورت دست نخورده دفع شود، اما کمک آن به گسترش ویروس هنوز ثابت نشده است.
2-5 انتشاردر مسافتهای دور
انتشار توسط انسان
شکی وجود ندارد که طی چند قرن گذشته، انسان مهمترین عامل گسترش بسیاری از ویروسها در سراسر جهان بوده است. در حالی که قبل از آن، این ویروسها به صورت محلی در یک یا تعداد کمی از نواحی جغرافیایی وجود داشته اند. این ویروسها در گیاه یا قطعات گیاهی و شاید بعضی مواقع از طریق ناقلین آنها، به وسیله سیب زمینی از آمریکا وارد اروپا شده و سپس از طریق غده به دیگر کشورها گسترش یافته اند.. ویروس موزاییک کاهو که به وسیله بذر انتقال مییابد، احتمالاً درهر جا که این محصول کشت شود، گسترش پیدا کرده است. این حقیقت که TMV میتواند به صورت واگیردار در سیگارهای تهیه شده از توتون بقا یابد، احتمالاً کافی است تا حدس بزنیم که این ویروس در تمام مناطق کشت اقتصادی توتون وجود دارد.
اثرات دیگری از فعالیتهای انسانی را میتوان از نظر تاریخی به دقت مشخص نمود. ویروس آبله آلو، در گونههای Prunus spp. ایجاد آلودگی نموده و به صورت ناپایا توسط شته سبز هلوM.persica و
Phorodon humuli انتقال مییابد.
شکل4 _ گسترش تدریجی پوتی ویروس آبله آلو (PPV ) در اروپا و آسیای غربی از سال 1915 به بعد. تاریخها بیانگر سالی میباشند که ویروس برای اولین باراز کشور گزارش شده است(Thresh,1984).
این بیماری، همانطور که در شکل4 نشان داده شده است، اولین بار در سال 1915 از بلغارستان گزارش شد و سپس از آنجا به اروپا گسترش پیدا کرد. شتهها، این ویروس را به باغات نزدیک گسترش داده اند ولی انتشار ویروس در مسافتهای طولانی، به احتمال زیاد نتیجه پخش مواد رویشی از گیاهان آلوده شده توسط انسان بوده است.
بیماری تریستزای مرکبات، اثر مخرب ویروس را زمانی که در مناطق جدید جایگزین میشود، نشان میدهد.ویروسها در مناطق جدید به سرعت منتشر میشوند، زیرا در این نواحی گیاهان میزبان ممکن است مقاومت اندکی داشته باشند و یا اصلاً مقاومت طبیعی در برابر ویروس تازه وارد به منطقه وجود نداشته باشد. کلستروویروس تریستزای مرکبات، قابل انتقال با شته، احتمالاً از آسیای جنوب شرقی نشأت گرفته و همراه موادگیاهی به آفریقای جنوبی و استرالیا وارد شده است. پس از ورود این ویروس به آمریکای جنوبی، با نواحی وسیعی از میزبان حساس و شته ناقل مؤثر، بین سالهای 1927 و 1930 میلیونها درخت پرتقال شیرین روی پایههای پرتقال ترش حساس از بین رفتند. سپس خسارات در ایالات متحده آمریکا، اسپانیا و اسرائیل نیز به وجود آمدند. در سالهای اخیر، چندین ویروس در انستیتوی ایستگاههای اصلاح گیاهی، در گیاهان حاصل از مواد گیاهی آلوده و در مواد اصلاحی پیدا شده اند. این ویروسها اغلب از آنجا به گیاهان زراعی اطراف، یا در بذر لینههای اصلاحی مورد آزمایش به محلهای دورتر و سپس به مواد تکثیری گیاهی، که برای فروش تکثیر میشوند، راه یافته اند. این امر، دلیل انتشار جهانی پوتی ویروس موزائیک بذر زاد نخود( PSbMV ) در حدود سال 1960 میباشد.
استابس در سال 1956 خاطر نشان کرد که ویروس عامل بیماری کوتولگی ابلق هویج در استرالیا که شته ناقل آن Cavariella aegopodii به فراوانی وجود دارد و فاقد دشمن طبیعی است، بسیار سریع انتشار مییابد. در مقابل، انتشار آن در کالیفرنیا به سبب پارازیته شدن ناقل توسط دشمن طبیعی، به کندی صورت میگیرد.
بطورکلی فعالیتهای انسانی ممکن است منجر به انتشار ویروس و ناقل در داخل کشور شود. برای مثال خاکهای قلمستانها در جنوب انگلیس غالباً شامل نپوویروسها و ناقلین آنها میباشند، که میتوانند همراه با گیاه میزبان، به خصوص در صورت نهفتگی بیماری ، به مناطق دیگر حمل شوند. در عین حال، ناقلین زنده ویروسها ممکن است همراه با وسائط نقلیه مانند هواپیمای مسافربر
ی و باربری ( بویژه مواد گیاهی مانند گیاهان زینتی) وارد شده و پس از آن، در محیطهای جدید ممکن است جایگزین شوند. تریپس گل غربی Frankliniella occidentalis ابتدا در غرب آمریکا یافت شد. این تریپس در دهه 1970 به شرق کوههای راکی تا به ایالات متحده شمالی، کانادا و به چندین کشور دیگر مانندهاوائی ، کره جنوبی ، نیوزلند و پرو گسترش یافت.در دهه 1980 این تریپس، به احتمال زیاد همراه با گیاهان زینتی، به چندین کشور اروپای شمالی وارد شد که در آنجا در گلخانه استقرار یافته و به جنوب اروپاکه این حشره قبلاً به فضای باز راه یافته، وارد شده است.گسترش این حشره منجر به اپیدمی فوق العاده چند میزبانه توسپوویروس پژمردگی لکه ای گوجه فرنگی (TSWV ) شده که در حال حاضر به صورت اپیدمیک در چندین گیاه زراعی در آمده است. بیوتیپ جدیدB و بسیار مهاجم سفید بالک پنبه که در اصل گرمسیری است گاهی اوقات به علت تولید عارضه برگ نقره ای در گیاهان تیره کدوئیان، یک گونه مختلف به نام Bemisia argantifolia در نظر گرفته میشود. بیوتیپ B عمدتاً جایگزین سفید بالک پنبه معمولی (بیوتیپA ) در جنوب غربی ایالات متحده آمریکا تا سال 1991 شده بود. این منجر به تغییر در بروز نسبی ویروسهای خانواده Closteroviridae و Geminiviridae منتقل شونده با مگس سفید، شد. بیوتیپ B در اوائل دهه 1990 ، احتمالاً همراه با گیاهان زینتی وارد اروپا شد. هم اکنون این بیوتیپ تقریباً در سراسر جهان، از جمله مناطق نیمه گرمسیری و معتدله، گسترش یافته است. اخیراً این حشره در اروپا به اپیدمیهای متعدد از جمله بیماریهای زردی در تعدادی از سبزیجات در گلخانهها و اطراف آنها کمک کرده است.
ناقلین هوازاد
شتههای ناقل برای انتشار ویروس در مسافتهای کوتاه و طولانی اهمیت دارند. این موضوع همان قدر که برای ویروسهای ناپایا مهم است، برای ویروسهای پایا نیز میتواند اهمیت داشته باشد. برای مثال، مطالعات مفصلی روی حرکت شتهها در انگلستان انجام گرفته است. این مطالعات نشان میدهد که در صورت وجود گیاهان میزبان مناسب، هر ساله کل این جزیره توسط بسیاری از گونههای مهم شته کلونیزه میشود. این حرکت مستلزم طی مسافتی بیش از هزار کیلومتر است که معمولاً با یک پرواز انجام نمی گیرد. در این شرایط ممکن است چندین کلونیزاسیون متوالی انجام گردد. انتقال ناپایا توسط شتهها به ندرت از 50 تا 150 متر تجاوز میکند.شپشکهای آردآلود که بادناویروس تورم جوانه کاکائو (CSSV ) را منتقل میکنند، ممکن است تنها در مسافتهای کوتاه بخزند و از درختی به درخت دیگر، که شاخههای آنها به یکدیگر پیچیده است، حرکت کنند. چنین انتشار شعاعی ویروس گاهی اوقات به همراه پخش جهشی توسط شپشکهای آردآلود بادزاد میباشد که منجر به شیوع ماهواره ای ( ازهم جدا ) در درههای کاکائو کاری شده است.شتهها و بویژه زنجرکهای برگی ممکن است برای چندین ساعت پرواز کنند. پس از به پرواز در آمدن، این حشرات ممکن است با باد به جریانات هوائی ارتفاع بالا یا جریانات شدید ارتفاع پائین وزیده شوند
. برای نمونه، شتههای ناقل و لوتیوویروس کوتولگی زرد جو ( BYDV ) از ایالت تگزاس، کانزاس و اکلاهما در ایالات متحده در جهت شمال به آیووا یا حتی به مانیتوبا در کانادا حمل میشوند. اینها مسئول هجوم زود هنگام ویروس کوتولگی زرد جو میباشند. به طور مشابه، نانوویروس کوتولگی شبدر زیرزمینی ( SCSV ) که حبوبات علفی و همچنین حبوبات غذائی یکساله را آلوده میکند، ممکن است مسافتهای طولانی را با ناقل ابرمهاجر خودAphis craccivora در استرالیا بپیماید.طول این مسافت از روی دریا دو هزار کیلومتر میباشد.
کنههای منتقل کننده ویروس، مانند Aceria tulipa که تریتیموویروس موزاییک
رگه ای گندم (WSMV ) را به طریق پایا منتقل مینماید، با باد تا فواصلی منتقل
می شوند(شکل 7 ).این انتقال در مسافتهای طولانی میسر نمی باشد برای اینکه کنه بسیار ظریف بوده و به سرعت خشک میشود. ویروسهای ناپایا به ندرت در مسافتهای دور قابل انتقال هستند. با وجود این، مدارک فزاینده ای وجود دارند مبنی بر اینکه ویروسهای منتقل شونده ناپایا ممکن است به مدت ساعتها، زمانی که مثلاً در حین پرواز در مسافتهای طولانی دسترس فوری به گیاهان ندارند، توسط شتهها نگه داشته بشوند. برای مثال، پوتی ویروس موزاییک کوتولگی ذرت ( MDMV ) به مدت 21 ساعت در شته Schizaphis graminum باقی میماند. این ویروس برای سالیان دراز در آمریکای جنوبی محدود میشد، اما یک اپیدمی بزرگ ناگهانی در ذرت شیرین در ایالت مینوستا و سایر ایالات شمالی در سال 1977، که با یک الگوی هوائی در ارتباط بود، امکان حمل غیر معمولی شته در مسافت طولانی از جنوب به شمال را فراهم آورد.
انتشار آبزاد
ویروسهای گیاهی آلوده کننده ای که از رودخانهها و دریاچهها جدا شدند شامل توباموویروسها، یک Potexvirus ، TNV ، STNV ، ویروس پیسک میخک، CMV و ویروس لکه حلقوی میخک میباشند. تعداد زیادی از این ویروسها بسیار پایدار بوده و فاقد ناقلین هوازادی هستند که گسترش آنها را در مسافتهای طولانی میسر سازد. این ویروسها در گیاهان، غلظتهای بالایی داشته و با آزاد شدن از ریشههای آلوده شده میتوانند بدون وجود ناقل، در ریشه گیاهان دیگر آلودگی ایجاد نمایند.
بسیاری از این ویروسها دامنه میزبانی وسیعی دارند. تعداد زیادی از ویروسهای آلوده کننده ، احتمالاً در نتیجه جذب روی سطح ذرات کلوئیدی آلی و غیر آلی به خصوص ذرات رس، به همراه آنها در آب حرکت میکنند. در این حالت آنها در مقایسه با ویروسهای آزاد، مقاومت بسیار بیشتری در برابر غیر فعال شدن از خود نشان میدهند. همچنین مشخص شده است که بعضی از این ویروسها میتوانند بدون از دست دادن توانایی آلوده سازی، از لوله گوارش انسان عبور کنند. شواهد دیگری نشان میدهد که فاضلابها میتوانند منبعی برای بعضی از ویروسهای آبزاد محسوب شوند. ویروسهای آبزاد ممکن است یکی از عوامل وقوع زوال جنگلها در اروپا باشند ولی برای روشن شدن پاسخ این سؤال، مطالعات بیشتری لازم دارد.
3) منابع آلودگی
آلودگی به ویروس اکثراً از گیاهان کشت شده به عنوان گیاهان زراعی یا از رویش طبیعی گیاهان وحشی، به وجود میآید. گیاهانی که از آنها حمله ویروس به گیاه محصول یا سایر جمعیتهای گیاهی شروع میشود، منابع اولیه آلودگی میباشند.زمانی که گیاهان جدیدآلوده میشوند، بلافاصله به صورت منابع ثانوی گسترش بعدی ویروس عمل میکنند و به همین ترتیب ادامه مییابد. مؤثر بودن گیاهان به عنوان منابع آلودگی بستگی به سرعت و اندازه تکثیر ویروس در آنها دارد. البته ممکن است گیاهانی که به عنوان منابع آلودگی عمل میکنند، همزمان به عنوان منابع آلودگی برای ناقل نیز(چنانچه ناقلین بر روی آنها تکثیر یابند) عمل کنند.
3-1 گیاهان زراعی
آلودگی اغلب از درون خود گیاه محصول نشأت میگیرد بویژه اگر گیاه زراعی از مواد رویشی آلوده به مواد تکثیری میکروارگانیسمهای آلوده کننده تولید شوند. چنین احتمال آلودگی برای مواد رویشی گیاهی معمولاً وجود دارد، اما در بذر برخی گیاهان محصول نیز ممکن است این احتمال بالا باشد. گیاهان حاصله از مواد رویشی آلوده به صورت بالقوه منابع اولیه وخطرناک آلودگی هستند برای این
که این گیاهان از همان آغاز رویش در میان گیاهان محصول حضور دارند. ویروسهای موزاییک پیسک سبز خیار ( CGMMV)، پیسک ملایم فلفل (PMMV ) وتوبموویروس موزاییک گوجه فرنگی (ToMV ) به صورت مکانیکی به آسانی گسترش بیشتری مییابند. پوتی ویروسهای موزاییک معمولی لوبیا (BCMV )، موزاییک کاهو (LMV )و موزاییک سویا (SMV )به آسانی از آلودگیهای بذر زاد توسط گونههای متنوع شته به طور ناپایا به سرعت گسترش پیدا میکنند. این پوتی ویروسها دارای طیف میزبانی محدود میباشند و بذر آلوده به آنها، منبع اولیه آلودگی است. بوجود آمدن اپیدمی ح
مله ویروسی در گیاهان دائمی مانند شبدر، رازک، مو و درختان میوه از احتمال نسبتاً بالایی برخوردار است برای اینکه وقتی گیاهان آلوده شدند ممکن است برای مدتهای طولانی به صورت منابع آلودگی عمل کنند.
آلودگی در گیاهان یکساله نیز ممکن است از گیاهان محافظ زمین یا داوطلب، یا ار گیاهان تکثیر شونده به طریق رویشی با حفظ دائمی آلودگی، یا از گیاهان حاصل از ریزش قبل از رسیدگی بذر در زمان برداشت، در صورتی که ویروسهاتوسط بذر قابل انتقال باشند، به وجود آید. باقی ماندن غدههای سیب زمینی در زمین در نتیجه برداشت مکانیزه، باعث نگرانیهای قابل ملاحظه ای در اروپای غربی پس از زمستانهای ملایم میشود. ریشههای مو در خاک تاکستان ممکن است برای چندین سال به زندگی خود ادامه دهند که در این مدت نماتدها می توانند نپوویروس برگ بادبزنی(GFLV ) را از آنها منتقل سازند. گیاهان رازک ممکن است از تکههای باقی مانده در زمین پس از برداشت و حفظ بقاء خود به مدت چندین سال در گندمیان علفی یا علفهای هرز زمین آیشی، و یا در گیاهان کشت شده بعدی، سبز شوند. نماتدها ممکن است نژاد رازک نپوویروس موزاییک آرابیس(ArMV ) را از آنها گرفته و موجب بیماری سرگزیدگی در گیاهان جدید رازک بشوند. گزارش شده است که انجام آبیاری میتواند جوانه زنی بقایای محصول مانند ریشهها یا کلش باقی مانده در زمین از گیاهان نظیر ریشههای نیشکر، پنبه، و برنج را افزایش داده و به این ترتیب به انتقال ویروسها و ناقلین آنها کمک کند.
گیاهان زراعی دائمی در مزارع مختلف اغلب با یکدیگر همپوشانی دارند. در آن صورت، آلودگی مزارع تازه کشت شده ممکن است از همان گونههای گیاهی موجود در مزارع مجاور نشأت بگیرد. فصلهای رویش گیاهان یکساله مانند چغندر، کلم و هویج که به صورت طبیعی همپوشانی ندارند ممکن است توسط گیاهانی که به منظور تولید بذری در همان ناحیه تولید میشوند، به هم ارتباط داده شوند. گیاهان زراعی یکساله با فصل رویش کوتاه نیز ممکن است در نواحی با زمستانهای ملایم که گیاهان زراعی به رشد خود ادامه میدهند یا به صورت سالم در مزرعه برای برداشت در بهار بعدی میمانند، همپوشانی داشته باشند. این امر در مورد تره فرنگی در اروپای غربی و چغندرقند در کالیفرنیا در فاصله بین سالهای 1960 و 1970 اتفاق افتاد که منجر به اپیدمیهای ویروسی شدند. ریسک اپیدمی ویروسی در اثر اصلاح برای مقاومت زمستانی یا هنگامی که کشت و داشت در ماههای سرد در تونلهای پلاستیکی یا ساختمانهای دیگر ارزان قیمت ادامه مییابد، افزایش بیشتری پیدا میکند. در شرایط آب و هوایی خشک، آبیاری ممکن است کشت محصول دوم را فراهم کند. این امر در مال نیجریه در سال 1975 منجر به اپیدمیهای بیسابقه بیماری پرپشتی بادام زمینی در محصولات دیمی بادام زمینی شد. بادام زمینیها از گیاهان آبیاری شده در فصل خشک قبلی یا از گیاهان داوطلب حاصله، آلوده شدند.
شناسائی منابع آلودگی در مزارع مختلف یک نوع گیاه زراعی موقعی که کولتیوارهای کشت شده به صورت غیرعادی به ویروس عکس العمل نشان میدهند یا حاملها فاقد علایم مشخص هستند ممکن است دشوار باشد. این مشکل در سیب زمینیريا، که کولتیوارها اغلب به صورت واضح به آلودگی ویروسی واکنش نشان نمی دهند، آشکار است، که با وجود نهفتگی ، کولتیوارهای حساس همجوار را تهدید میکنند.
آلوده شدن گیاهان زراعی اغلب از گونههای گیاهی کاملاً متفاوت صورت میگیرد. پوتی ویروس موزاییک زرد لوبیا (BYMV )، که عمدتاً میروس حبوبات میباشد، ممکن است برخی گیاهان دیگر را آلوده سازد. اکثر گلایولهای زینتی آلودگی ویروسی دارند، اگرچه از ویروس رنج نمی برند و این گیاهان در صورتی که در جوار لوبیا کاشته شوند ممکن است تهدید جدی به لوبیای معمولی معمولی باشند. اخیراً معلوم شده است که لوبیا به عنوان میزبان رابط بگوموویروس پیچیدگی برگ زرد گوجه فرنگی ( TYLCV ) در گیاهان گوجه فرنگی عمل میکند. ویروسهای دیگر دارای طیفهای میزبانی وسیعی هستند. به عنوان نمونه، کوکوموویروس موزاییک خیار (CMV ) حدود 800 گونه گیاهی مشتمل بر گیاهان چوبی، متعلق به حدود 85 خانواده گیاهی را میتواند آلوده کند. گیاهان منبع آلودگی ممکن است کاملاً فاقد علامت ابتلا به ویروس باشند. یونجه آلوده به ویروس برگ قاشقی لوبیا در تابستان یا در کشورهای گرمسیری علایمی نشان نمی دهد یا ممکن است تنها زردی رگبرگ در بخش خنک فصل، به صورتی که در اروپای غربی مشهود است، نشان دهد. این ویروس و ویروسهای دیگر خانواده Luteoviridae به صورت پایا توسط در مسافتهای طولانی انتقال مییابند. این نوع ویروسها موجب زردی شدید و کوتولگی در تعدادی از حبوبات یکساله که غذای
دام هستند، مانند لوبیاها( شامل لوبیای معمولی و باقلا)، نخود ایرانی، لوبیا چشم بلبلی، عدس، باقلای مصری یا لوبیا گرگی، نخود و سویا و در لگومهای مناسب برای تغذیه دام میشوند.سطوح قدیمی شبدر اغلب طیف وسیعی از ویروسها، از جمله پوتی ویروس موزاییک زرد لوبیا ( BYMV )، کوکوموویروس کوتولگی بادام زمینی (PSV ) ، نپوویروس لکه حلقوی توتون ( TRSV )و پوتکس ویروس موزاییک شبدر سفید (WClMV ) را در خود پناه میدهند.
